Komplekse tilpasninger og neo-darwinistisk matematikk
(Fritt utvalgt fra Darwins Doubt; S.C.Meyer, Harper One, 2013, Ch.12.A) {med oversetters kommentar i klamme-parentes}
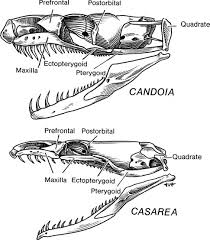 Biolog Tom Frazzetta kjente neo-darwinistisk teori så godt som noen: Evolusjonære mekanismer forandrer organismer gradvis, med modifiserte økninger, oppdelt i form av kontinuerlige endringer, der en strukturell tilstand glir gradvis over i en annen. (1) Imidlertid hadde Frazzetta sine tvil. Som en ekspert på funksjonelle biologiske mekanismer, som studerte hvordan dyr faktisk fungerte. Han hadde dissekert hodeskaller av sjeldne slanger, som bare var funnet på Mauritium i det Indiske hav. Disse slangene, bolyeriner -er boa-lignende, men har en anatomisk egenskap som ikke er funnet i andre virveldyr. Deres øvre-kjevebein er delt i to segmenter, som er linket av en fleksibel sammenføyning, og betjent av mange ekstra nerver, vev og ulikt arrangerte leddbånd. Dette unike trekket tillater slangen å bøye den fremre halvdel bakover når de angriper bytte. Bilde 1. Nederste Casarea er av type bolyerine
Biolog Tom Frazzetta kjente neo-darwinistisk teori så godt som noen: Evolusjonære mekanismer forandrer organismer gradvis, med modifiserte økninger, oppdelt i form av kontinuerlige endringer, der en strukturell tilstand glir gradvis over i en annen. (1) Imidlertid hadde Frazzetta sine tvil. Som en ekspert på funksjonelle biologiske mekanismer, som studerte hvordan dyr faktisk fungerte. Han hadde dissekert hodeskaller av sjeldne slanger, som bare var funnet på Mauritium i det Indiske hav. Disse slangene, bolyeriner -er boa-lignende, men har en anatomisk egenskap som ikke er funnet i andre virveldyr. Deres øvre-kjevebein er delt i to segmenter, som er linket av en fleksibel sammenføyning, og betjent av mange ekstra nerver, vev og ulikt arrangerte leddbånd. Dette unike trekket tillater slangen å bøye den fremre halvdel bakover når de angriper bytte. Bilde 1. Nederste Casarea er av type bolyerine
Kunne dette komplekse systemet av ben, ledd, vev og leddbånd ha utviklet seg gradvis? En bevegelig sammenføyning, som deler kjevebenet i to segmenter," bemerket Frazzetta , "synes å ha enten nærvær eller fravær, med ingen mellomledd til å forbinde de to tilstandene." (2) "Hvordan kan et kjevebein være halvveis delt?" Siden mellomformene ikke ville være virksomme/fungerende, ville det å lage en bolyerin-kjeve innebære at alle nødvendige deler måtte oppstå samtidig. Likevel gikk problemet for neo-darwinismen langt utover anatomiske eiendommeligheter ved sjeldne slanger. Som ung professor i evolusjonsbiologi, hadde han studert komplekse egenskaper i en stor variasjon av arter. Han visste at nesten enhver biologisk struktur av interesse, det indre øret, øyne, gjeller, lunger, fjær, reproduksjonsorganer, kretsløp og åndedretts-system, innehadde flerfoldige nødvendige komponenter.
 Å endre et slikt system, krever å endre hvert av de uavhengige delene som deres funksjonalitet er basert på. Det kan ikke skje på måfå. F.eks. endrer man ett av de tre benene i pattedyrs indre øre, så vil det kreve korresponderende endringer i de andre bena, og også i andre deler av øret, som i membranen eller sneglehuset. Komplekse biologiske systemer avhenger ofte av hundrevis av slike uavhengige, men likevel samvirkende, nødvendige deler. I det antallet av nødvendige deler øker, øker antall nødvendige, koordinerte endringer også, -forutsatt at virkemåten skal bevares. Og om ikke, så vil naturlig seleksjon sile ut disse ikke-fungerende endringene. Vansken med å vedlikeholde funksjonell integritet for systemet, mens en modifiserer én av delene, øker stadig. Det kan sammenlignes, i følge Frazzetta med 'gradvis forbedring av en maskin, mens den er i drift!' (5)
Å endre et slikt system, krever å endre hvert av de uavhengige delene som deres funksjonalitet er basert på. Det kan ikke skje på måfå. F.eks. endrer man ett av de tre benene i pattedyrs indre øre, så vil det kreve korresponderende endringer i de andre bena, og også i andre deler av øret, som i membranen eller sneglehuset. Komplekse biologiske systemer avhenger ofte av hundrevis av slike uavhengige, men likevel samvirkende, nødvendige deler. I det antallet av nødvendige deler øker, øker antall nødvendige, koordinerte endringer også, -forutsatt at virkemåten skal bevares. Og om ikke, så vil naturlig seleksjon sile ut disse ikke-fungerende endringene. Vansken med å vedlikeholde funksjonell integritet for systemet, mens en modifiserer én av delene, øker stadig. Det kan sammenlignes, i følge Frazzetta med 'gradvis forbedring av en maskin, mens den er i drift!' (5)
Bilde 2. Advarsel mot slikt
Historisk har evolusjonsbiologer forsøkt å løse dette problemet med én fordelaktig variasjon eller mutasjon, om gangen. Denne fremgangsmåten startet med Darwin selv, som ba sine lesere 'forestille seg en serie av fordelaktige endringer, i form av en nerve som er sensitiv for lys.' (6) Selv om biologer på Frazzettas tid, som nå, ikke fullt ut forstår hvordan DNA kan korrelere eller avbilde høyere ordens, komplekse morfologiske strukturer, så visste de også da (1975) at hundrevis av gener kunne være involvert i å kode for en enkelt, kompleks integrert struktur. Å endre anatomisk struktur av pattedyrs øre eller virveldyrs øyne, f.eks., ville innebære å endre genene som kodet for deres bestanddeler. Det innebar igjen at høyst usannsynlige, multiple koordinerte mutasjoner ville måtte inntreffe nærmest samtidig.
Frazzetta forklarte: "Fenotype endringer av integrerte systemet krever et usannsynlig sammentreff av genetiske, og herav arvelige fenotype endringer, av et tett, spesifisert slag (8). Genetiske endringer som påvirker enhver av de nødvendige delene, må matches med mange korresponderende og koordinerte endringer, om de ikke skal ende i funksjonstap, evt. død." Derfor konkluderte Frazzetta: "Vi er enda overlatt det aldri minkende behov for å forklare evolusjonære endringer i systemer, som har operasjonelle integrasjons-karakteristika, i likhet med ting vi gjenkjenner som maskiner." (9) På den tiden fikk det liten tiltrekningskraft, da en regnet med en bortimot ubegrenset kreativ kraft ved mutasjoner og naturlig seleksjon.
Bilde 3. Fra animasjon om DNA-kopiering
Som støtte for sitt syn, knyttet de seg til matematiske uttrykk for neo-darwinistisk teori i populasjons-genetikken. Populasjonsgenetiske modeller viser hvordan gen-frekvenser endrer seg som følge av mutasjons-prosesser, genetisk drift (nøytrale endringer i genomet i forhold til seleksjon) og naturlig seleksjon. Disse modellene ble laget under formodningen at fordelaktige variasjoner eller trekk kan oppstå som følge av én enkelt mutasjon. Ut fra dette beskrives hvor mye evolusjonær endring som kan oppstå i en tidsperiode. Tre faktorer er grunnlag for estimatene: mutasjonsrater, effektive populasjons-størrelser og dannelses tid. Når evolusjonsbiologer setter inn estimater for disse tre variable, synes beregningene å vise, at standard evolusjonære mekanismer kan iverksette signifikante evolusjonære endringer i mange slags organismer. Endog det å bygge komplekse systemer. Så lenge mutasjoner generer en kontinuerlig forsyning av nye trekk. så kan ethvert system bli bygd trekk for trekk. Det sier i hvert fall teorien..
Tillit til disse matematiske modellene og deres underliggende formodninger, fører til at mange neo-darwinister ikke ser behov for å gi detaljerte redegjørelser for evolusjonære stier, som komplekse systemer har oppstått gjennom. F.eks sies dette av biologene Paul Ehrlich og Richard Holm: "En trenger ikke gå i detalj omkring evolusjon av fuglens vinge, giraffens nakke eller virveldyrets øye.. i det de utvalgte opphav til disse og andre strukturer for atferds-mønstre kan bli antatt å være i hovedsak det samme som for industriell melanisme.. Selv en liten fordel eller ulempe i en spesiell endring, forsyner en tilstrekkelig forskjell for virkningen til naturlig seleksjon." (10) Her viser ordbruken 'tilstrekkelig forskjell' til en av faktorene i populasjons-genetikk, den såkalte seleksjons-koeffisienten, som bestemmer hvor raskt et trekk gjennomtrenger populasjonen. Beskjeden er klar: "matematikken forteller historien, biologiske detaljer omkring opprinnelsen til komplekse systemer betyr ikke noe.
Mange av datidens evolusjonsbiologer, slik som grunnleggerne av populasjons-genetikk, antok at det allerede eksisterte en mekanisme for å bygge nye gener. Faktisk antok de at nye trekk, og genene som bygde dem, kunne oppstå som resultat av enkeltmutasjoner, evt. en serie mutasjoner, der hver bidro med en liten økt fordel. Men hva om det finnes systemer i levende organismer, som ikke kan bli bygd én mutasjon om gangen, og i stedet må bygges ved simultane, koordinerte endringer? Hva om å bygge et enkelt gen eller protein, krever slike koordinerte mutasjonsendringer? Hva om individuelle gener viste seg å være komplekse tilpasninger?
Det har lenge vært tendensen i evolusjonsbiologien å betrakte matematiske utfordringer, á la Axe sin utfordring om hvor sjeldne fungerende proteiner er, som noe utenfra, -fra bio-ingeniører eller statistikere, og dermed som noe eksotisk eller irrelevant. Dette har begynt å endre seg. I senere tiår har utvikling i molekylærbiologien og populasjons-genetikk, avslørt en sammenheng mellom problemet med opprinnelsen til nye gener, og opprinnelsen til komplekse sammenføyninger. Det startet med Frazzettas tvil i 1975.
 Populasjonsgenetikk og opprinnelsen til genetisk informasjon
Populasjonsgenetikk og opprinnelsen til genetisk informasjon
Likningene til populasjonsgenetikerne ble dannet ut fra 1930-tallets forutsetninger om at enkelt-mutasjoner kunne endre ett gen slik at det kunne produsere ett nytt trekk. Etter oppdagelsen av DNA-strukturen i 1953, så endret oppfatningen av mutasjoner seg. Nå ble de mer sett på som enkelt 'trykkfeil' i tusenvis av nukleotide-sekvenser. Da ble det mer som en stavefeil i en lengre tekst. I stedet begynte mange vitenskapsfolk å se at individuelle mutasjoner, i stedet for å danne nye fordelaktige trekk, ville de mye mer sannsynlig degradere eksisterende trekk. Likeså at akkumulering av mutasjoner til slutt ville ende i funksjonstap. Dermed kom det nye teorier på 1970-tallet for hvordan mutasjoner kunne danne nye gener, med idéene om gen-duplisering, påfølgende nøytral evolusjon og positiv seleksjon. Etter teorien var formulert, tenkte mange evolusjonsbiologer at det var oppdaget en mekanisme som sikret evt. produksjon av nye gener, uten å kompromittere virkemåten til organismen.
Så da Frazzetta konfronterte evolusjonsbiologien med problemet med komplekse tilpasninger midt på 1970-tallet, ble han møtt av ett 'kollektivt gjesp'. Populasjonsgenetikkens matematiske modeller impliserte at det ikke var noen utfordring, dersom formodningene om hvorvidt mutasjoner kunne danne nye trekk, holdt stikk. Men var de det? Kunne en serie av mutasjoner generere nye gener for å danne nye proteiner og karakteristika, eller krevde det multiple, koordinerte mutasjoner?
Bilde 4. Opprinnelsen til genetisk informasjon?
Er gener komplekse tilpasninger?
Klassiske darwinistiske evolusjonsbiologer har forutsatt at små, separate, steg-for-steg endringer, kunne produsere alle nødvendige biologiske strukturer og karakteristika, forutsatt at hver endring innebar en overlevelses- eller reproduksjons-fordel. I sitt kap. om 'Darwin og moderne vitenskap' så beskrev den britiske genetikeren W. Bateson tørt hvor vidtgående denne formodningen var, og at den hindret evolusjonærbiologer å konfrontere den virkelige vanskeligheten, ved å forklare opprinnelsen til komplekse tilpasninger: "Ved å foreslå at stegene til en tilpasset mekanisme er ubestemte og ikke-sensible, så er alt videre bryderi spart. Mens det kunne påstås at artene oppsto ved ubestemte og ikke-sensible mekanismer, så var det ikke behov for å slite oss ut ved å prøve å forstå prosessene. Dette arbeidsbesparende rådet ble funnet gunstig". (12)
En av de første prominente evolusjons-biologene som betraktet muligheten at nye gener og proteiner kunne kreve multiple, koordinerte mutasjoner var den selverklært 'overbeviste darwinisten' John M. Smith. Han arbeidet som fly-ingeniør under 2.verdenskrig, men tok så opp studiet av evolusjonsbiologi etter krigen. I 1970 besvarte han en artikkel av Frank Salisbury, som stilte spørsmål ved evnen tilfeldige mutasjoner hadde til å kunne forklare spesifisiteten ved nukleotidene, for å danne fungerende proteiner. I følge Salisburys beregninger, ville den mutasjonelle mekanismen komme til kort med flere hundre 10-er potenser, i løpet av 4 milliarder år, selv for ett enkelt nødvendig gen. M.Smith foreslo en modell for protein evolusjon. En serie av små gradvise forbedringer, der hver sekvens ivaretok sin funksjon underveis. Smith satte opp en pedagogisk skisse av hvordan det var tenkt: WORD-WORE-GORE-GONE-GENE. Hvor ordene skulle symbolisere proteiner, og bokstavene aminosyrer. Hvert steg i utviklingen skulle være fra ett fungerende protein til ett annet. (15)
Likevel var M. Smith betenkt for seg selv: Hvordan kunne ett gen eller protein utvikle seg til ett annet, dersom det krevde flerfoldige, simultane endringer i arrangementet av aminosyrer? Om bygging av nye gener krever multiple koordinerte mutasjoner, så vile sannsynligheten for å danne ett nytt gen eller protein, synke dramatisk, siden en slik endring ikke bare ville kreve én usannsynlig begivenhet, men kanskje to, tre eller flere.. Her er hvordan han beskrev det potensielle problemet:
Anta ett protein ABDCD ...eksisterer, og at proteinet abCD ville være fordelaktig mht. seleksjon. Anta videre at mellomledd som aBCD og AbCD ..er ikke-funksjonelle. Disse formene ville dannes ved mutasjoner, men vil vanligvis elimineres ved naturlig seleksjon, før en annen mutasjon kan inntreffe. Det doble steget fra ABCD .. til abCD, ville dermed bare svært usannsynlig kunne inntreffe (17). I følge M. Smiths syn, utgjorde usannsynligheten i tilknytning til doble eller multiple steg av koordinert mutasjon, et signifikant potensielt problem for  molekylær evolusjon. Imidlertid konkluderte han til slutt med at slike mutasjoner var så usannsynlige, at de 'måtte ikke' ha spilt noen viktig rolle i evolusjonen av nye strukturer." Han forklarte, "slike doble steg .. kan av og til inntreffe, men er sannsynligvis for sjeldne til å ha spilt en viktig rolle i evolusjon." (18) I flere tiår var problemet han luftet forsvunnet fra oppmerksomheten. I 2005 påpekte biokjemiker H. Allen Orr i Nature Reviews Genetics: "Selv om Maynard Smiths arbeid dukket opp tidlig i den molekylære revolusjonen, så ble hans idéer om å betrakte protein-evolusjon glemt i nesten to tiår." (19) Ikke før i det 21. århundre konfronterte biologer utfordringen om å gjøre rigorøse kvantitative analyser av sannsynlighet, for protein-til-protein evolusjon.
molekylær evolusjon. Imidlertid konkluderte han til slutt med at slike mutasjoner var så usannsynlige, at de 'måtte ikke' ha spilt noen viktig rolle i evolusjonen av nye strukturer." Han forklarte, "slike doble steg .. kan av og til inntreffe, men er sannsynligvis for sjeldne til å ha spilt en viktig rolle i evolusjon." (18) I flere tiår var problemet han luftet forsvunnet fra oppmerksomheten. I 2005 påpekte biokjemiker H. Allen Orr i Nature Reviews Genetics: "Selv om Maynard Smiths arbeid dukket opp tidlig i den molekylære revolusjonen, så ble hans idéer om å betrakte protein-evolusjon glemt i nesten to tiår." (19) Ikke før i det 21. århundre konfronterte biologer utfordringen om å gjøre rigorøse kvantitative analyser av sannsynlighet, for protein-til-protein evolusjon.
Å vente på komplekse tilpasninger
Biokjemiker Michael Behe, ved Lehigh University introduserte i 2004 sammen med fysiker David Snoke ved Phittsburg universitet, en artikkel i journalen Protein Science, som tok opp problemet John M. Smith hadde introdusert. (20) Behe hadde på den tiden presentert seg som en prominent kritiker av neo-darwinismen. Han hevdet at dens mekanismer ikke ga fullgod forklaring på funksjonelt integrert "ikke-reduserbar kompleks" molekylære maskiner. Han og Snoke forsøkte å fastsette sannsynligheten til protein-evolusjon i det tilfelle den faktisk krevde flere koordinerte mutasjoner. De benyttet analyse-modeller fra populasjonsgenetikk, for å gjøre evalueringen. Det var den standard evolusjonære modellen, der en ser for seg gener som oppstår gjennom gen-duplisering, og påfølgende mutasjoner i den dupliserte genen.
Bilde 5. Def. av molekylære maskiner
Behe og Snoke fastsatte sannsynligheten ved denne modellen for multi-cellulære organismer, i det tilfellet at multiple (to eller flere) punktmutasjoner måtte inntreffe samtidig for å danne ett nytt selekterbart gen eller protein. Der M. Smith så behovet for multiple koordinerte mutasjoner som et potensielt problem, så argumenterte Behe og Snoke for at evolusjonære biologer trengte å vurdere det, og de kvantifiserte alvoret ved saken.
De bemerket først at mange proteiner, som et resultat av sin funksjon, krevde unike kombinasjoner av aminosyrer, som virket på koordinerte måter. Så henviste de til en autoritativ lærebok, Molecular Evolution, av evolusjonær biolog Wen-Hsiung Li. Der beskriver Li at det å utvikle molekyl-bindene kapasitet i proteiner, slik som hemoglobin, kan kreve mange mutasjons-steg, (21) selv om det første stegene på veien til å danne slik kapasitet ikke vil tilføre noen selektiv fordel. Som Li erklærer, "å oppnå en ny funksjon kan kreve mange mutasjons-steg, og et poeng som trenger å understrekes er at de første stegene kan være selektivt nøytrale, fordi den nye funksjonen ikke er manifestert før et visst antall steg allerede har inntruffet." (22).
 På grunn av dette, mente Behe og Snoke at en serie av atskilte mutasjoner ikke kunne generere molekyl-bindings funksjon i et protein, som ikke tidligere hadde denne egenskapen. Det trengtes i stedet multiple, koordinerte mutasjoner. Behe og Snoke gjorde tilsvarende vurderinger for utvikling av protein til protein-interaksjoner. De bemerker at for at proteiner skal samhandle med hverandre på spesifikke måter, må typisk minst flerfoldige, individuelt nødvendige aminosyrer være til stede i kombinasjon i hvert protein, som igjen, indikerer behovet for multiple koordinerte mutasjoner.
På grunn av dette, mente Behe og Snoke at en serie av atskilte mutasjoner ikke kunne generere molekyl-bindings funksjon i et protein, som ikke tidligere hadde denne egenskapen. Det trengtes i stedet multiple, koordinerte mutasjoner. Behe og Snoke gjorde tilsvarende vurderinger for utvikling av protein til protein-interaksjoner. De bemerker at for at proteiner skal samhandle med hverandre på spesifikke måter, må typisk minst flerfoldige, individuelt nødvendige aminosyrer være til stede i kombinasjon i hvert protein, som igjen, indikerer behovet for multiple koordinerte mutasjoner.
Så mange endringer på liten tid
Behe og Snoke benyttet prinsippene fra populasjonsgenetikk, for å fastsette sannsynligheten til ulike antall koordinerte mutasjoner i en gitt tidsperiode. De spurte: "Er det mulig at det var nok tid i evolusjonær historie til å generere koordinerte mutasjoner? Om så var, hvor mange koordinerte mutasjoner er det sannsynlig å forvente i en gitt tidsperiode, gitt ulike populasjons-størrelser, mutasjonsrater og generasjonsvarighet? Så for ulike kombinasjoner av disse variable beregnet de hvor lang tid det ville ta å frambringe tre eller mer koordinerte mutasjoner. Det de fant ut generelt, var at sannsynligheten for multiple mutasjoner, som oppsto i nære funksjonelt relevante, koordinasjoner, var "avskrekkende" (prohibitively) liten. Det ville sannsynligvis ta ekstremt lang tid, typisk mye lenger enn alderen på jorda.
Bilde 6. Eks. på motorproteiner
Referanser:
1. Frazzetta, "From Hopeful Monsters to Bolyerine Snakes?" s.62-63
2. Frazzetta, "From Hopeful Monsters to Bolyerine Snakes?" s.63
5. Frazzetta, "Complex Adaptations in Evolving Populations, s.20
6. Som Darwin skrev i 'Origin': Om vi må sammenligne øyet med et optisk instrument, kan vi forestille oss vi tar et tykt lag av gjennomsiktig vev, med en lys-sensitiv nerve, og så forestille oss at hvert lag kontinuerlig endres sakte i tetthet, for å skille lag med ulik tetthet og tykkhet, og så med overflaten til hvert lag sakte endring av form." s.188-189
8. Frazzetta, "Modeling Complex Morphological Change in Evolution," s.129
9. Frazzetta, "Modeling Complex Morphological Change in Evolution," s.130
10. Ehrlich and Holm, The Process of Evolution, s.157
12. Bateson, "Heredity and Variation in Modern Lights," s.83-84
15. Maynard Smith, "Natural Selection and the Process of Protein Space," s.564
17. M. Smith, "Natural Selection and the Process of Protein Space," s.564
18. "Natural Selection and the Process of Protein Space," s.564
19. Orr, "The Generic Theory of Adaptation," s.123
20. Behe og Snooke, "Simulating Evolution by Gene Duplication of Protein Features that Require Multiple Amino Acid Residues"
21. Wen-Hsiung, Molecular Evolution, s.427
22. Wen-Hsiung, Molecular Evolution, s.427
Stoffutvalg og bilder ved Asbjørn E. Lund